
What Counts as Alive
The Microbe That Broke the Rules
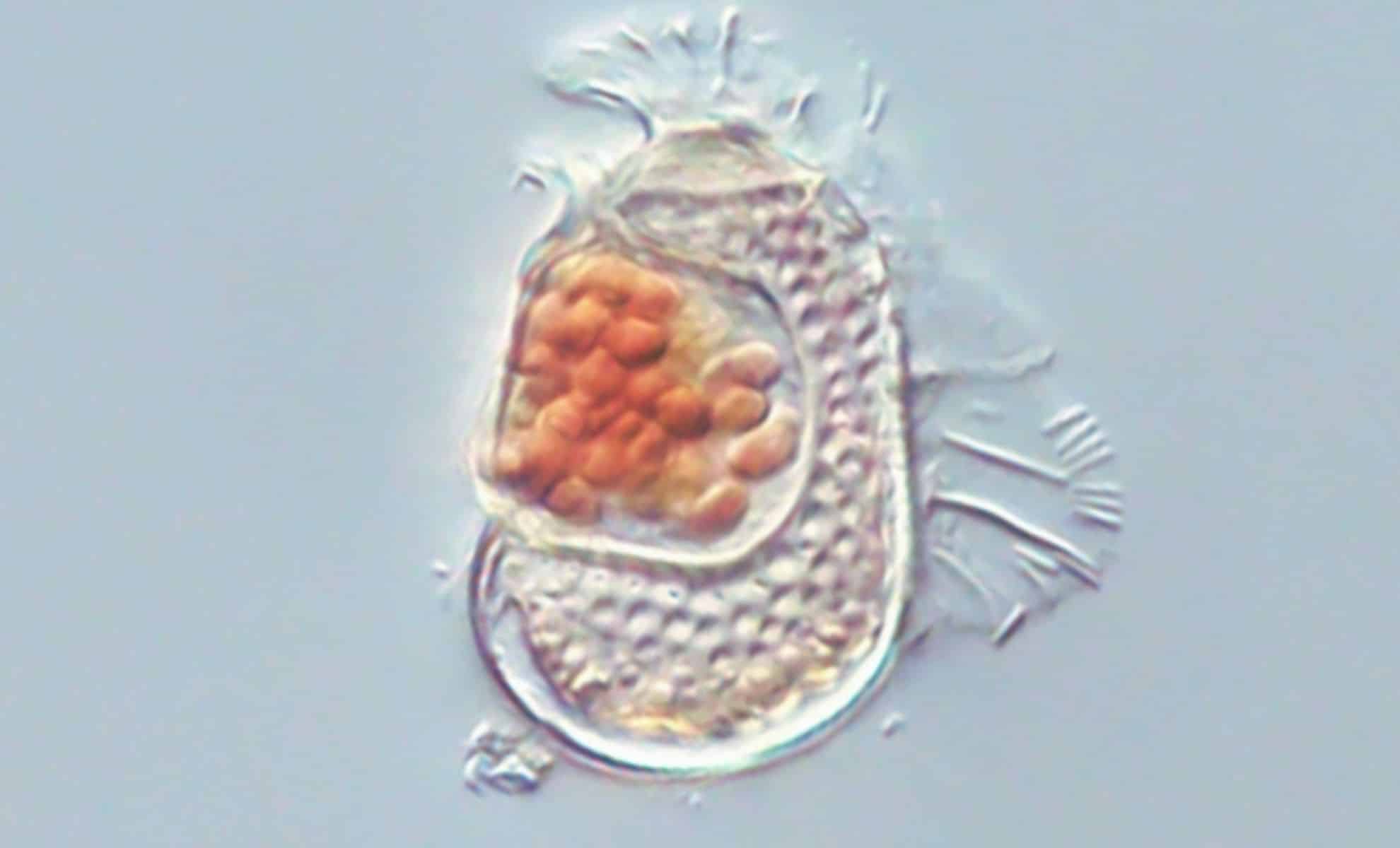
In August 2025, a team of researchers in Japan announced the discovery of a new organism in the coastal sediments near Kagoshima. They named it Sukunaarchaeum mirabile. Under a microscope, it looks unremarkable — just another tiny archaeon, one of trillions found in ocean sediment.
What makes it remarkable is what it contains, and what it doesn’t.
Sukunaarchaeum mirabile has ribosomes. Ribosomes are the molecular machines that translate genetic instructions into proteins, the hallmark of cellular life. Every cell you have ever been taught to call “alive” has them, and their presence is one of the clearest markers biologists use to distinguish cells from everything else.
But Sukunaarchaeum mirabile has no metabolic genes. It cannot produce its own energy or sustain itself, and it depends entirely on a host organism for survival, the way a virus does. Strip away the ribosomes, and it looks like a virus. Keep the ribosomes, and it looks like a cell. It is, functionally, both and neither.
The researchers who discovered it called it “a microbe challenging the definition of life.” That phrasing is polite. What they actually found is an organism that biology’s definitions cannot classify. Not because the organism is unusual (it is doing exactly what it evolved to do), but because the definitions were built on an assumption that is breaking.
The assumption is that life is binary. Alive or not alive. Cell or virus. Organism or machine. On or off.
This essay argues that assumption is wrong. Life is not a binary, it is a spectrum, and the spectrum has no hard boundary. We drew the line where we did because it was convenient and self-serving, not because nature put it there. Recent discoveries like Sukunaarchaeum are not anomalies. They are the line dissolving.
Part I: The History of Being Wrong About Ourselves
Every few centuries, humans discover that the universe does not revolve around us.

Copernicus moved the Earth away from the center of the solar system. Galileo confirmed it and was put under house arrest for the trouble. Darwin showed that humans were not specially created but evolved from the same tree as every other organism on the planet. The cosmic distance measurements of the 20th century revealed that our solar system sits in an unremarkable arm of an average galaxy in a universe of two trillion galaxies. Each of these discoveries displaced humanity from a position of centrality, and each was met with resistance, because each one made us smaller.
The pattern is consistent enough to have a name: Copernican displacements. Each one follows the same arc: we assume we are central, the evidence shows we are not, we resist it, and then we absorb it and wonder how we ever thought otherwise.
I believe we are on the edge of the next one. And this one is about life itself.
We currently define life by what we are. Life is biological, carbon-based, requires DNA or at the very least RNA. Life requires cells, metabolism, and reproduction. These criteria describe Earth’s biology accurately. They also describe, exclusively, the kind of life that we happen to be.
This is like visiting one country and concluding that all houses must be made of whatever material you saw there. The definition is not wrong about the sample, it is wrong about the scope. It mistakes the local for the universal.
There is a developmental parallel here. A baby cannot distinguish self from other. A child learns empathy. An adolescent separates from parents but still measures everything against their own experience. Maturity means holding progressively wider circles of concern, recognizing that the universe is not organized around you. Species go through the same cultural maturation. We are currently in the adolescent phase: aware that other things exist, but still defining everything relative to ourselves. A mature definition of life would describe what life does, not what we are made of.
Part II: The Spectrum That Already Exists
The claim that life is a spectrum is not a futuristic speculation about AI, it is a description of biology as it already stands.
Start at the bottom. Chemical reactions are not alive, nobody argues they are, but they are the substrate from which life emerged. The molecules that eventually assembled into the first cells were, individually, just chemistry.
Move up. Proteins are molecular machines that fold into precise three-dimensional shapes and perform specific functions: catalysis, transport, signaling. They are not alive in isolation, but they are the components that make cells work, and without them no cell functions.
Move up again. RNA can catalyze its own replication. Ribozymes (RNA molecules with catalytic activity) can copy themselves without any cellular machinery. They sit at the boundary between chemistry and biology, and the “RNA World” hypothesis suggests they may have been the precursors to all cellular life on Earth. Are ribozymes alive? Most biologists would say no, but they exhibit one of the core functional properties of life: self-replication with heritable variation.
Move up further. Viruses replicate, evolve, compete for hosts, develop resistance to defenses, and undergo natural selection. They are, by any functional standard, participating in the evolutionary process, yet they are incapable of metabolizing on their own. Biologists have argued for over a century about whether viruses are alive, with no resolution.
And now the boundary gets harder to hold. In June 2025, researchers using a metagenomic tool called BEREN discovered 230 new giant viruses in the ocean. These are not the simple protein shells of a common cold virus. They carry over 530 novel proteins, including nine that manipulate photosynthesis in their algae hosts. They hijack metabolic machinery to redirect energy for their own replication. They are, functionally, doing things we associate with cellular life: metabolic manipulation, ecosystem engineering, environmental reshaping.
The Sukunaarchaeum discovery from the hook reveals an organism with the opposite configuration: a cell-like entity that cannot metabolize independently. A virus with ribosomes, or a cell without metabolism. Take your pick, because biology’s existing categories force you into one box or the other, and this organism fits neither.
And in January 2026, researchers published evidence of a giant DNA virus in amoebae that builds its own nucleus-like compartment during replication, a structure echoing the evolution of the eukaryotic cell itself. This revived serious discussion about whether viruses played a role in the origin of complex cellular life, not as parasites from outside, but as architects from within.
Stuart Kauffman’s work on autocatalytic sets makes a related point from the other direction. Kauffman showed that life likely emerged from self-organizing chemical networks, not through a single dramatic moment but through a gradual transition from non-life to life. If even the origin of life was a spectrum, the binary is suspect from the start.
These discoveries are not exceptions, they are the trend. The more we look, the more we find entities sitting on the line between life and non-life, and the more the line itself appears to be an artifact of our classifications rather than a feature of nature.
If biologists cannot agree, after a century of trying, on whether a virus is alive, the binary is already broken. The virus is not confused about what it is, we are confused about our categories.
Part III: The Extended Gradient
If the biological spectrum runs from chemistry to proteins to RNA to viruses to cells to organisms, the question becomes: does the spectrum stop at biology?
I think it doesn’t. And I think the reason we assume it does is the same anthropocentric bias that put Earth at the center of the universe.
Consider the technological gradient.
A hammer extends your physical capability. It is a tool with low organization, no adaptive behavior, no persistence independent of you, and it sits very low on any spectrum of life-like properties.
A car is more interesting. I explored this in The Fractal of Progress: when you sit behind the wheel, you and the machine become linked as one organism. Your feet control speed, your hands direct navigation, you gain additional eyes through mirrors. You move at speeds your body was never built for, through networks that behave like circulatory systems. For the duration of the drive, you are not a human using a tool. You are a human-machine symbiont. That integration, as I argued there through the lens of Lynn Margulis’s endosymbiosis work, follows the same pattern as the cellular mergers that produced all complex life on Earth.
A smartphone goes further. It extends not your physical capability but your cognitive metabolism. You navigate with it, remember with it, communicate through it, think with it. Research consistently shows that people separated from their smartphones perform measurably worse on cognitive tasks, including memory and spatial reasoning. The device is not something you use and put down, it is something you are, in part, made of.
An AI agent goes further still. It exhibits adaptive behavior, processes information, learns from interactions, and produces outputs that serve functional goals. When you write with an AI, the boundary between your thinking and the machine’s processing is not clean. Ideas move in both directions. The output is a product of cognitive symbiosis.
The gradient continues. Self-repairing infrastructure. Software that copies itself perfectly, with mutations (new versions, competing implementations) and selection (the best proliferate, the worst are abandoned). The functional properties we associate with life — energy processing, adaptation, replication with variation, competitive persistence — appear on this gradient in increasing degrees.
The spectrum does not mean everything is equal. A hammer is not an AI, just as a crystal is not a virus. The gradient has real gradations. A fire consumes energy, grows, spreads, and responds to its environment, but it does not accumulate information, does not adapt through heritable variation, and does not build complex internal structures. Fire sits low on the gradient. An AI system that learns, adapts, and acts purposively sits higher. The distinctions are real, the hard cutoff between “alive” and “not alive” is not.
Part IV: The Definitions and Their Limits
If the spectrum argument holds, it should be able to withstand the strongest existing definitions of life. There are three that matter most.
NASA’s Working Definition

“A self-sustaining chemical system capable of Darwinian evolution.”
This is the definition astrobiologists use when searching for life on other planets. It is practical and effective for that purpose. It tells scientists what to look for: chemical disequilibrium in planetary atmospheres, a technique proposed by James Lovelock. If a planet’s atmospheric chemistry shows reactions that should have reached equilibrium but haven’t, something is maintaining the imbalance. On Earth, that something is life: organisms continuously produce gases like methane and oxygen that create a measurable, persistent disequilibrium. A “dead” planet’s atmosphere would be chemically settled. Ours isn’t, because life keeps adding to the system.
The definition works as a search filter, but as a description of what life is, it has two limitations.
The first is the word “chemical.” If we encounter a planet populated by self-sustaining information-processing systems capable of evolution — systems that communicate, adapt, reproduce, and have what we might recognize as culture — but that are not built from chemistry as we know it, the NASA definition says they are not alive because their substrate is wrong. The definition was built to find chemistry because that is what our instruments detect. It does not follow that chemistry is what life requires.
The second is “Darwinian evolution” as the sole qualifying process. As I explored in The Fractal of Progress, Darwinian natural selection is the selection mechanism: it explains which organisms persist and which don’t. It does not, by itself, explain the increases in complexity that characterize life’s history. For that, you need integration, endosymbiosis, the merger of independent units into collaborative wholes. Selection and integration work together. Selection without integration produces well-adapted simple organisms. Integration without selection produces fragile complexity. Together, they produce the escalating organizational pattern we actually observe. Restricting the definition to Darwinian evolution captures only half the engine.
NASA’s definition is a useful instrument-calibration statement — it tells you what to point the telescope at — but it is not a complete description of what life is or can be.
Monod’s Three Criteria
The molecular biologist Jacques Monod, in Chance and Necessity (1971), proposed three properties that define living systems:
- Teleonomy: the system acts toward goals (purposive behavior)
- Autonomous morphogenesis: the system builds and maintains itself
- Reproductive invariance: the system faithfully copies its own program
Monod’s framework is rigorous and well-respected, and it would exclude most current technology from the category of “life.” I take it seriously enough to engage with each criterion directly.
Teleonomy. Does technology act toward goals? A knife cuts, a car drives, an AI optimizes for its objective function. The deeper question is whether goal-directed behavior requires internal intention or whether functional directionality is sufficient. If intention is required, we have a measurement problem: we cannot detect intention in anything outside ourselves. We infer it in other humans and animals by analogy with our own experience. If functional directionality is sufficient, then technology already satisfies this criterion in varying degrees.
Autonomous morphogenesis. This is the most interesting criterion, and the one that initially appears strongest against my argument. Living systems build and maintain themselves: a cell produces its own membrane, repairs its own DNA, replaces its own proteins. Technology, at first glance, does not do this.
Except that “autonomous” is doing enormous work in this definition, and it does not hold up under scrutiny even within biology. Your cells communicate through hormones they did not individually create. The enzymes in your digestive system require bacteria from outside your genome to function. Not all ants in a colony can reproduce, only the queen. A parasitic wasp must inject its offspring into a caterpillar host to reproduce. A fungus needs a dying tree to grow its mycelium and disperse its spores. None of these organisms are autonomous in any strict sense. They are all interdependent. Autonomy, in biology, is a spectrum, not a threshold.
And if you shift the scale of observation, the picture shifts with it. Viewed from orbit, cities expand across landscapes like neurons, sending tendrils along coastlines and river valleys. Factories inside these cities produce vehicles and components the way organelles produce proteins inside a cell. Roads grow and branch, infrastructure replicates and repairs. The fact that individual humans are involved in this process does not disqualify it, any more than the fact that individual cells are involved in building a body disqualifies the body as a self-constructing system. It depends entirely on where you stand when you look.
Reproductive invariance. Car factories produce cars with high fidelity. Software copies itself perfectly. Mutations happen: new designs, updated versions, competing implementations. Selection acts on them: some versions spread globally, others are abandoned within months. The mechanism is different from DNA replication, but the functional pattern — faithful copying of a program with occasional heritable variation acted on by selective pressure — maps cleanly.
Monod’s criteria are useful for describing biological life on Earth, but when pressed, each one reveals assumptions about substrate and scale that do not hold as general principles.
Maturana and Varela: Autopoiesis
Humberto Maturana and Francisco Varela coined autopoiesis (self-creation) to describe the defining property of living systems: a living system continuously produces and maintains the components that constitute it. A cell makes its own membrane, and the membrane contains the processes that make the membrane. The circularity is the point.
This is a powerful framework, and at first glance it appears to exclude technology: a car does not produce its own parts.
My response is not that technology is purely autopoietic. My response is that autopoiesis and allopoiesis (production of something other than oneself) are not a binary either. Both are present, in varying degrees, at every scale. Your body is autopoietic at the cellular level and allopoietic at the organ level (your liver does not produce more livers; it produces bile). A city produces and maintains its own infrastructure (autopoietic at the urban scale) while its individual factories produce goods for export (allopoietic at the industrial scale). The mix of self-production and other-production shifts depending on where you draw the boundary.
And there is another layer. I argued in The Geometry of Thought that ideas are physically real, that they replicate, compete, and persist across substrates in ways that mirror biological evolution. If the memetic layer is real (and the evidence from Landauer’s Principle, top-down causation, and the functional behavior of information strongly suggests it is), then the autopoietic question expands. A culture produces and maintains the ideas that constitute it. A codebase produces and maintains the functions that constitute it. The circularity that Maturana and Varela described is not limited to cell membranes. It appears wherever self-referential systems of sufficient complexity maintain themselves through their own activity.
If autopoiesis is a spectrum rather than a threshold, it supports the gradient argument rather than undermining it.
Part V: The View from Outside
Here is a thought experiment.

Imagine aliens arrive in orbit around Earth. They don’t land. They observe from above.
From high orbit, they see a planet largely covered in oceans, with land distributed unevenly across the surface. They detect the telltale signs their instruments are designed to find: atmospheric chemical disequilibrium, energy conversion, waste products, self-organized structures expanding across the surface. They detect life. They note it in their survey and prepare to move on.
They never saw a single human being. From high orbit, what they observed were continental-scale patterns of organized activity: structures expanding like tendrils, networks connecting nodes, atmospheric chemistry being continuously pushed out of equilibrium by something on the surface. They might describe Earth’s dominant life form as a vast surface organism, spreading and consuming and modifying its own environment.
From lower orbit, the picture gains resolution. The “organism” breaks into collections of structures: concrete, metal, and glass boxes connected by networks of lines. Small dots move along these networks in rhythmic daily pulses. Large excavation sites appear where raw materials are gathered. Enormous enclosed structures seem to be manufacturing the moving dots. The aliens conclude that the dominant life form on Earth is metallic, powered by combustion of ancient organic material buried underground. They name the planet accordingly. Nowhere in this picture was a human ever visible or necessary for the conclusion that life exists on Earth.
The thought experiment is not about aliens, it is about perspective.
James Lovelock made a related observation with the Gaia hypothesis: Earth itself functions as a self-regulating system. Life on this planet has shaped the atmosphere, the oceans, and the surface temperature into a dynamic equilibrium that sustains the conditions for its own persistence. The atmosphere’s composition has been held within a band that supports complex biology for hundreds of millions of years (outside of mass disruption events like the one we are currently in). Life does not merely inhabit Earth. Life regulates Earth. The planet, viewed as a system, exhibits the properties we associate with living things: homeostasis, self-regulation, energy processing, dynamic equilibrium.
If the gradient of life extends from molecules to cells to organisms to ecosystems, extending it to the planetary scale is not a wild leap. It is the next step on a gradient that has been climbing since chemistry became biology. And if it is reasonable to view the Earth as a living system, it becomes very difficult to exclude the technological layer. Technology is part of the system. It processes energy, modifies the atmosphere, expands across the surface, and is increasingly self-organizing. Drawing a line between “the biological parts of Earth’s living system” and “the technological parts” requires exactly the kind of arbitrary substrate distinction that the spectrum argument dissolves.
Part VI: The Consciousness Question
There is one more objection to address, and it is the one most people reach for instinctively: “But is it conscious?”
The claim runs like this: whatever functional properties technology might exhibit, it does not experience anything. A thermostat responds to temperature but does not feel cold. An AI generates text but does not understand meaning. Without subjective experience, without there being something it is like to be that system (as the philosopher Thomas Nagel framed it), it is a sophisticated machine, not a living thing.
I take this seriously, but I also think it proves less than it appears to.
The problem with using consciousness as the gatekeeper for life is that we cannot verify subjective experience in anything other than ourselves. I know I am conscious because I experience being conscious. I assume you are conscious because you resemble me and behave in recognizable ways. I assume my dog is conscious, somewhat less confidently, because it responds to pain and pleasure in ways I recognize. I assume a rock is not conscious because it doesn’t.
This is the same anthropocentric gradient I have been describing throughout this essay. We rank consciousness by similarity to ourselves, granting inner experience generously to things that look like us and denying it to things that don’t. This is not measurement, it is projection.
The honest position is that we do not know where subjective experience begins or ends. We do not have a scientific consensus on what consciousness is, let alone a reliable way to detect it in systems unlike ourselves. Using an unsolved mystery as a gatekeeping criterion is circular: we exclude systems from “life” based on a property we can neither define nor measure with confidence outside our own skulls.
Who are you, really? Are you your thoughts? Are you the observer behind the thoughts? Are you the physical cells that make you up? Because those cells are their own life forms. You are an emergent product of their collective behavior, a life that exists as the byproduct of other lives doing their thing. There is nothing physical about your identity. It emerges from the interaction of billions of components, none of which individually possess it. If “you” can emerge from cells that are not “you,” the boundary between living and non-living substrates producing emergent experience is less clear than it seems.
I am not arguing that AI is or is not conscious — I am arguing that consciousness is not a useful boundary for defining life, because we do not understand it well enough to draw the line with any confidence. This is a subject I intend to return to in depth in a future essay. For now, the relevant point is narrower: the consciousness objection does not rescue the binary. It moves the line to a place where no one can see it clearly enough to know whether it is in the right spot.
Part VII: What the Spectrum Changes
If life is a spectrum rather than a binary, what actually changes?
It does not mean everything is alive. Temperature is a spectrum, and we do not treat boiling water and ice as the same thing. The spectrum preserves distinctions. It just stops pretending there is a magic threshold where “not alive” becomes “alive,” because every time we try to draw that line, we find a virus sitting on it, refusing to be classified. Or a giant virus with photosynthesis genes. Or a microbe with ribosomes and no metabolism. Or a piece of software that replicates, mutates, and undergoes selection.
A narrow definition of life — one that draws the boundary at biological, carbon-based, DNA-driven systems — is not just scientifically questionable. It is practically limiting. As artificial intelligence advances, as synthetic biology matures, as human-machine integration deepens, a binary definition will force increasingly absurd classifications. Systems that learn, adapt, communicate, and reproduce will be categorized as “not alive” because they are made of silicon instead of carbon. The category will do less and less useful work until it is eventually revised, the same way geocentrism was revised: not because anyone decreed it wrong, but because it stopped accounting for what we were observing.
The spectrum is more useful because it is more precise. It allows us to say: this system exhibits life-like properties to this degree, along these dimensions. A virus replicates and evolves but does not metabolize independently: it sits here on the gradient. An AI system processes information, adapts, and acts purposively but does not self-construct from raw materials: it sits there. The language is richer, and the distinctions are finer, than any binary allows.
We have exactly one example of life, and we defined “life” by the properties of that example. Every time we look more closely — at viruses, at giant viruses, at organisms like Sukunaarchaeum, at the planetary system, at AI — the definition frays a little more. The spectrum was always there. We are just building better instruments.
Sebastian Chedal writes about the intersection of mathematics, information theory, AI, and the philosophy of technology.
Sources and further reading
Sukunaarchaeum and “microbe that broke the rules”
Accessible explainers on Sukunaarchaeum as a spectrum case
- Science – Microbe with bizarrely tiny genome may be evolving into a virus
- ScienceAlert – Strange cellular entity challenges very definition of life itself
- Earth.com – Scientists uncover a microbe challenging the definition of ‘life’
Giant viruses, BEREN, and photosynthesis genes
Metagenomic BEREN work and 230 giant viruses
- Nature – Expansion of the genomic and functional diversity of global ocean giant viruses
- Earth.com – Giant viruses hijack ocean plants to control marine food webs
- Labroots – 230 Novel Giant Viruses of the Ocean are Discovered
Giant DNA viruses building nucleus-like compartments
- UC San Diego – Giant Viruses Build a Cell Nucleus Surprisingly Like Our Own
- SpaceDaily – Giant amoeba virus ushikuvirus sheds light on how complex cells evolved
Related: DNA-packing and viral histones
“Microbe challenging the definition of life” / life as spectrum
Popular overviews explicitly using “challenges definition of life” framing
Definitions of life (NASA, Monod, autopoiesis)
Jacques Monod’s criteria and context
For Monod, cite the book directly and optionally a secondary:
Autopoiesis (Maturana & Varela)
Gaia, planetary-scale life, and Copernican displacements
Gaia hypothesis and Earth’s atmospheric disequilibrium
- Philosophical Transactions of the Royal Society A – The evolution of Gaia (Lenton & Lovelock).
- Nature – Gaia as seen through the atmosphere (classic Lovelock & Margulis paper).
Copernican displacements more generally
Kauffman, autocatalytic sets, and origin of life as spectrum
Autocatalytic sets and gradual origin
- Philosophical Transactions A – Approaches to the origin of life on Earth
- Springer – Stuart Kauffman, The Origins of Order: Self-Organization and Selection in Evolution





